میتوز
 میتوز
میتوز
مرحله میتوز تقسیم یاختهای نسبت به جنبههای دیگر چرخه یاختهای یوکاریوتها توجه بیشتری را به خود جلب کرده است. بر اساس سنتی که وجود داردمیتوز را به چهار مرحله پروفاز ، متافاز ، آنافاز و تلوفاز تقسیم میکنند. این گونه تقسیم بندی آسان به نظر میرسد، اما روند تقسیم حقیقتا پیوسته است و مراحل تقسیم به آرامی از یک مرحله به مرحله دیگر وارد میشود.
را به چهار مرحله پروفاز ، متافاز ، آنافاز و تلوفاز تقسیم میکنند. این گونه تقسیم بندی آسان به نظر میرسد، اما روند تقسیم حقیقتا پیوسته است و مراحل تقسیم به آرامی از یک مرحله به مرحله دیگر وارد میشود.
پروفاز
 اولین مرحله تقسیم یاختهای که با متراکم شدن کروموزومها (که آغاز آن از مرحله
اولین مرحله تقسیم یاختهای که با متراکم شدن کروموزومها (که آغاز آن از مرحله  است) آغاز میشود و شاید طولانیترین مرحله بوده و چند ساعت طول میکشد،پروفاز نام دارد. متراکم شدن کروموزومها در طی مرحله پروفاز ادامه مییابد. بنابراین کروموزومهایی که در آغاز پروفاز به صورت رشتهای ظریف بودند، در آخر این مرحله کاملا حجیم میشوند. هنگامی که بخشی از کروموزوم که حاوی ژنهای RNA ریبوزومی است، متراکم میشود، سنتز RNA ریبوزومی کاهش مییابد و در نتیجه هستک که قبلا مشخص شود، ناپدید میگردد.در حین متراکم شدن کروموزومها یک سری رویدادهای مهم دیگر رخ میدهد. هستک ناپدید شده و دستگاه ریزلولهای ، جهت جدا کردن کروموزومهای دختر تشکیل میگردد. در اوایل مرحله پروفاز دو جفت سانتریول از یکدیگر دور میشوند و بین آنها محوری از ریزلولهها تشکیل میگردد که رشتههای دوک نامیده میشوند. سانتریولها از هم دور میشوند تا در دو قطب مخالف روبروی هم قرار گیرند و پلی از ریزلولهها بین آنها بوجود میآید. در یاختههای گیاهی چنین پلی متشکل از رشتههای دوک بین قطبهای مخالف یاخته تشکیل میگردد، اما در این حالت مرکز سازمان دهنده ریزلولهها با میکروسکوپ نوری دیده نمیشود.
است) آغاز میشود و شاید طولانیترین مرحله بوده و چند ساعت طول میکشد،پروفاز نام دارد. متراکم شدن کروموزومها در طی مرحله پروفاز ادامه مییابد. بنابراین کروموزومهایی که در آغاز پروفاز به صورت رشتهای ظریف بودند، در آخر این مرحله کاملا حجیم میشوند. هنگامی که بخشی از کروموزوم که حاوی ژنهای RNA ریبوزومی است، متراکم میشود، سنتز RNA ریبوزومی کاهش مییابد و در نتیجه هستک که قبلا مشخص شود، ناپدید میگردد.در حین متراکم شدن کروموزومها یک سری رویدادهای مهم دیگر رخ میدهد. هستک ناپدید شده و دستگاه ریزلولهای ، جهت جدا کردن کروموزومهای دختر تشکیل میگردد. در اوایل مرحله پروفاز دو جفت سانتریول از یکدیگر دور میشوند و بین آنها محوری از ریزلولهها تشکیل میگردد که رشتههای دوک نامیده میشوند. سانتریولها از هم دور میشوند تا در دو قطب مخالف روبروی هم قرار گیرند و پلی از ریزلولهها بین آنها بوجود میآید. در یاختههای گیاهی چنین پلی متشکل از رشتههای دوک بین قطبهای مخالف یاخته تشکیل میگردد، اما در این حالت مرکز سازمان دهنده ریزلولهها با میکروسکوپ نوری دیده نمیشود.
 هنگام تشکیل دستگاه دوک ، غشای هسته خرد میشود و مواد آن بوسیله شبکه آندوپلاسمی جذب میگردد. بنابراین دوک ریزلولهای از یک قطب به قطب دیگر کشیده میشود. موقعیت دوک صفحه تقسیم را مشخص میکند که از مرکز هسته میگذرد و عمود بر دوک است. هنگامی که سانتریولهاییاختههای جانوری در حال تقسیم ، به قطبهای یاخته میرسند، در اطراف خود رشتههایی از ریزلولهها را به صورت شعاعی بیرون میفرستند که همانند داربستی سانتریولها را در برابر غشای یاخته نگاه میدارند. این نوع آرایش ریزلولهها را ستاره (آستر) میگویند.
هنگام تشکیل دستگاه دوک ، غشای هسته خرد میشود و مواد آن بوسیله شبکه آندوپلاسمی جذب میگردد. بنابراین دوک ریزلولهای از یک قطب به قطب دیگر کشیده میشود. موقعیت دوک صفحه تقسیم را مشخص میکند که از مرکز هسته میگذرد و عمود بر دوک است. هنگامی که سانتریولهاییاختههای جانوری در حال تقسیم ، به قطبهای یاخته میرسند، در اطراف خود رشتههایی از ریزلولهها را به صورت شعاعی بیرون میفرستند که همانند داربستی سانتریولها را در برابر غشای یاخته نگاه میدارند. این نوع آرایش ریزلولهها را ستاره (آستر) میگویند.
یاختههای گیاهی که دیواره سخت یاختهای دارند، فاقد سانتریول هستند. در حین ادامه پروفاز دومین گروه ریزلولهای از هر سانترومر منفرد به قطبهای دوک کشیده ممیشوند. از هر کروموزوم دو ریزلوله خارج میشود که دو طرف سانترومر را به دو قطب دوک متصل میکنند. دو ریزلوله متصل به سانترومر به پیشروی خود ادامه میدهند تا هر دو با دو قطب یاخته تماس حاصل کنند. این امر سبب کشیدگی کرومزومها به خط میانی یاخته میشود. در مرحله پروفاز ATP به مقدار فراوان مصرف میشود. اگر فهالیت تنفسی یاخته در ابتدای این مرحله متوقف گردد، میتوز هم متوقف خواهد شد. در اواخر این مرحله ، اگرچه یاخته مقدار زیادی ATP ذخیره میکند، با این وجود کاهش فعالیت تنفسی هیچ اثری بر میتوز ندارد.
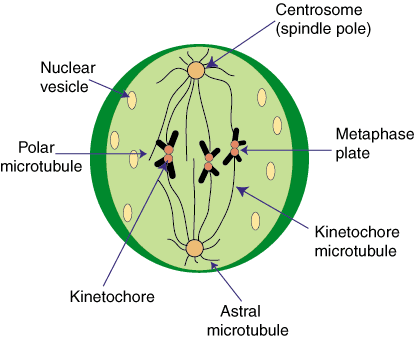
متافاز
دومین مرحله میتوز ، یعنی متافاز ، هنگامی آغاز میشود که کروماتیدهای جفت در مرکز یاخته در یک سطح قرار بگیرند. در این مرحله کروموزومها به آسانی قابل شمارش هستند. ریزلولهها که به دو طرف قطب کشیده شدهاند، آشکارا قابل مشاهدهاند. کروموزومها هم موازی یکدیگر نیستند، بلکه بازوی بلند آنها در هر جهتی قرار میگیرد. سانترومرها به یک فاصله از دو قطب در یک سطح قرار میگیرند.
در تقسیم میتوز کروموزومها ، سانترومرهای خود را بطور غیر فعال دنبال میکنند. هر سانترومر دو طرف دارد و یک ریزلوله سانترومری به هر طرف آن متصل شده و به قطبهای مخالف کشیده میشود. این نظم و ترتیب برای روند میتوز کاملا مهم است. هر اشتباهی در استقرار این ریزلولهها خطرناک است. به عنوان مثال ، اتصال دو ریزلوله سانترومری به همان قطب سبب جدا نشدن کروماتیدهای خواهر و در نتیجه باقی ماندن آنها در همان یاخته دختر میشود.
زیست شناسان برای متوقف کردن میتوز در مرحله متافاز ، از مادهای به نام کلشیسیناستفاده میکنند تا ساختار ریختی و تعداد کروموزومها را مطالعه کنند. در بسیاری از گونهها ، اندازه کروموزومها متغیر است. کروموزومهای بزرگتر در بیرون و کروموزومهای کوچکتر در مرکز قرار میگیرند. در این مرحله کروموزومها ضخیم و کوتاهاند. کرومونما به حداکثر انقباض خود میرسد. در پایان مرحله متافاز ، سانترومرها تقسیم میشوند. تقسیم سانترومر همه کروموزومها همزمان صورت میگیرد. شکل استوانهای کروموزومهای متافاز نتیجه محکم کلاف شدن کرومونماست. کوتاه شدن کروموزومهای پروفازی و تبدیل آنها به کروموزومهای متافازی هنگام تقسیم 3 تا 6 برابر است.
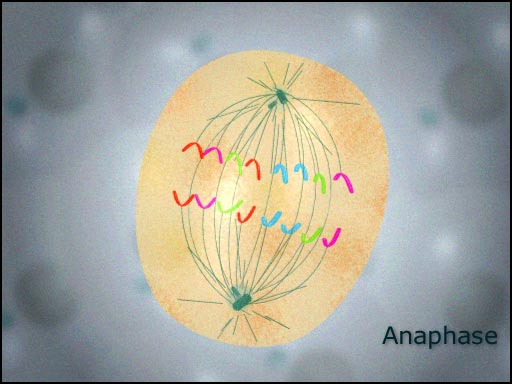
آنافاز
در بین مراحل میتوز ، ATPیرانده شده در اثر تغییرات ساختاری پروتئینهای پل ریزلولههای جفت است. چون هر یک از ریزلولههای جفت از نظر فیزیکی ، به قطبهای مخالف قلاب شدهاند، سر خوردن هر یک از آنها روی دیگری سبب دور شدن قطبها از یکدیگر میشود.سانترومرها به طرف قطبها حرکت میکنند. با جابجا شدن پیاپی اجزای توبولین از انتهاهای قطبی خود ریزلولههای سانترومری کوتاه میشوند. این روند کوتاه شدن ، یک انقباض نیست، چون ریزلولههای سانترومری مرکز تنظیم کننده جابجا میشوند. هر قدر اجزا توبولین بیشتر جابجا شوند، بی نظمی پیشرفته ریزلوله حامل کروموزوم سبب کوتاهتر شدن آن میگردد و کروموزوم را به قطب یاخته نزدیکتر میکشاند. رشتههای اکتین در دوک هم دیده میشوند.
تلوفاز
جدا شدن کروماتیدها که در مرحله آنافاز صورت میگیرد، تقسیم صحیح ژنوم همانندسازی شده را که عنصر اصلی میتوز است، کامل میکند. با این تکمیل ایفای نقش تلوفاز که از بین بردن تشکیلات دوک و تشکیل دو هسته است، آغاز میشود. دستگاه دوک متلاشی میشود. ریزلولهها نظم خود را از دست میدهند و به صورت مونرمرهای توبولین در میآیند و آماده استفاده مجدد در ساختار اسکلت یاختهای جدید میشوند.
غشای هسته در اطراف هر گروه کروماتیدهای دختر شکل میگیرد. این کروماتیدها که هنوز به شکل کروموزوماند، شروع به باز شدن میکنند و کاملا کشیده میشوند و به صورت کروماتین تجلی مییابند. یکی از ژنهایی که به سرعت ظاهر میشود، RNA ریبوزومی است که سبب ظهور مجدد هستک میگردد.

سندرم ترنر
بیماری هموفیلی
بیماری هموفیلی هموفیلی نوعی بیماری ارثی است که در آن خون در محل بریده شده بند نمیآید و جز در موارد استثنایی این بیماری فقط در افراد مذکر دیده میشود. عارضه خونریزی در هموفیلی میتواند بسته به شدت اختلال ژنتیکی درجات مختلفی از نظر شدت داشته باشد. |
انواع هموفیلی
هموفیلی A و B اختلالات انعقادی وابسته به کروموزوم جنسی هستند که به ترتیب بر اثر جهشهایی در ژنهای F8C و F9 ایجاد میشوند. جهشهای F8C موجب کمبود یا اختلال عملکرد عامل انعقادی 8 و جهشهای ژن F9 باعث کمبود یا اختلال عملکرد عامل انعقادی 11 میشوند. هموفیلی نوعی اختلال در تمام نژادها و بدون ترجیح نژادی است. میزان شیوع هموفیلی A از هموفیلی B بیشتر است.
شرح بیماری
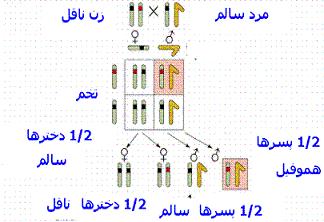
این بیماری از طریق مادر به پسرانش منتقل شده و مردان خوشبختانه نمیتوانند بیماری را به پسرانشان منتقل کنند. هموفیلی در 85 درصد موارد ناشی از کمبود فاکتور انعقاد خون شماره 8 است و این نوع هموفیلی موسوم به هموفیلی A یا هموفیلی کلاسیک است. در 15 درصد دیگر بیماران هموفیلیک ، تمایل به خونریزی بر اثر کمبود فاکتور انعقادی 11 بوجود میآید. هر دوی این فاکتورها بطور ژنتیکی از طریق کروموزوم X به صورت یک خاصیت مغلوب انتقال مییابند.
بنابراین تقریبا هیچگاه یک زن مبتلا به هموفیلی وجود نخواهد داشت زیرا لااقل یکی از کروموزومهای جنسی او دارای ژنهای سالم خواهند بود. اگر یکی از کروموزومهای جنسی زن معیوب باشد وی یک ناقل هموفیلی خواهد بود و این بیماری را به نیمی از فرزندان پسر خود منتقل خواهد کرد و حالت ناقل بیماری بودن را به نیمی از دختران خود انتقال خواهد داد.
علایم شایع
علایم اولیه هموفیلی خونریزیهای طولانی پس از خراشهای کوچک است. البته عقیده عمومی بر این است که اینگونه خونریزیهای کوچک ولی طولانی باعث مرگ شخص مبتلا نمیشود ولی به تدریج بیماری پیشرفت نموده علایم شدیدتری از قبیل خونریزیهای دردناک داخل مفصلی مانند مفصل زانو ایجاد خواهد کرد.
اینگونه حوادث با کوچکترین تحریکی پیش میآید و احتمالا یک پسر بچه را از انجام بازی محبوبش یعنی فوتبال و یا انجام هر کار کوچک دیگری که احتمال دارد زانو دچار پیچیدگی شود باز میدارد. تجمع خون در مفاصل ممکن است باعث خشک شدن مفصل شده و کودک را بطور کامل فلج کند و یا خونریزیهای غیر منتظرهای در ماهیچهها به وقوع بپیوندد.
فنوتیپ و سیر طبیعی
هموفیلی معمولا بیماری مردان است هر چند ندرتا خانمها هم به علت انحراف غیر فعال شدن کروموزوم ایکس از حالت طبیعی مبتلا میشوند. از نظر بالینی این دو نوع هموفیلی غیر قابل افتراق هستند. هر دوی اینها با خونریزی به داخل بافتهای نرم ، ماهیچهها و مفاصل متحمل وزن مشخص میشوند. خونریزی ظرف چند ساعت تا چند روز پس از تروما رخ میدهد و اغلب تا چند روز یا چند هفته ادامه مییابد.
آنهایی که بیماری شدیدی دارند معمولا در دوره نوزادی به علت هماتوم شدید سر یا خونریزی طولانی از زخمهای ناف یا محل ختنه تشخیص داده میشوند. افراد دچار بیماری متوسط اغلب تا زمانی که شروع به خزیدن یا راه رفتن نکنند دچار هماتوم نمیشوند و لذا تا آن زمان بیماری آنها تشخیص داده نمیشود.
درمان
اگر چه کار آزماییهای فعلی ژن درمانی بسیار امیدوار کننده به نظر میرسند، هیچ درمانی علاج دهندهای بجز پیوند کبد برای هموفیلی A و B وجود ندارد.
هر گاه شخص مبتلا به هموفیلی دچار خونریزی شدید و طولانی شود تقریبا تنها درمانی که واقعا موثر است تزریق فاکتور انعقادی شماره 8 خالص است. قیمت این فاکتور بسیار گران بوده و زیاد نیز در دسترس نیست زیرا اولا این فاکتور فقط میتواند از خون انسان و آنهم فقط به مقادیر فوقالعاده اندک بدست آورد. فاکتور 8 که به روش مهندسی ژنتیکتهیه شده به زودی برای مصرف انسانی در دسترس قرار خواهد گرفت.
خطر توارث
اگر خانمی سابقه خانوادگی هموفیلی داشته باشد با تجزیه و تحلیل پیوستگی یا شناسایی دو ژن جهش یافته F8C و F9 تجمع یافته در خانواده میتوان وضعیت حامل بودن او را تعیین کرد. تشخیص افراد حامل با سنجش آنزیمی دشوار میباشد و همه جا مقدور نیست. اگر مادری حامل باشد هر یک از پسرانش به احتمال 50 درصد ژنهای جهش یافته را به ارث خواهند برد. اگر مادری دارای پسری مبتلا به هموفیلی باشد اما هیچ خویشاوند مبتلای دیگری نداشته باشد احتمال حامل بودن او 2 در 3 است.
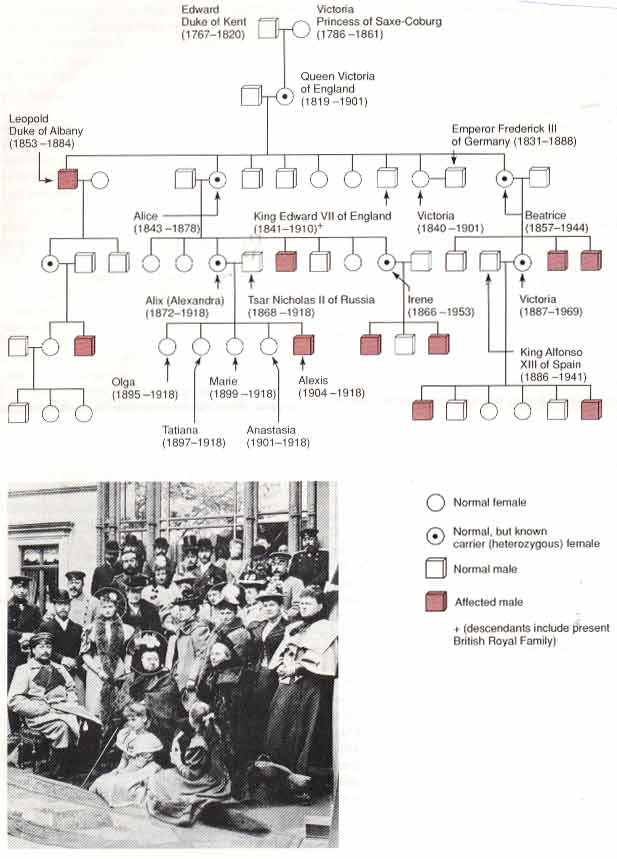
شجره نامه ی هموفیلی در خانواده ی ملکه ویکتوریا:
ویروس
سلولهای زنده همواره در فعالیت میباشد. چون ویروسها در خارج از بدن میزبان به حالت خنثی بسر میبرند به این مفهوم نمیتوان آنها را موجود زنده در نظر گرفت. معهذا هنگامی که ویروسها وارد سلول میزبان میشوند اسیدهای نوکلئیک آنها فعال گشته و منجر به تکثیر ویروس میگردد. از نظر بالینی ویروسها را میتوان موجودات زنده در نظر گرفت زیرا آنها مانند باکتریها ، قارچهای بیماریزا آلودگی و بیماری ایجاد میکنند. به ویروس کامل ویریون گفته میشود.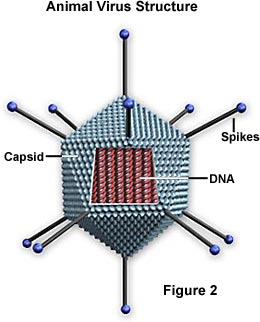
ساختمان شیمیایی ویروس
اسید نوکلئیک
تا 50 درصد در برخی از باکتریوفاژها متغیر است. برخلاف سلولهای پروکاریوتیک و یوکاریوتیک که همواره دارای DNA به عنوان ماده ژنتیکی اصلی خود هستند ویروسها دارای یکی از دو نوع اسید نوکلئیک بوده و هرگز هر دو را باهم ندارد. اسید نوکلئیک در بعضی ویروسها به شکل خطی و در بعضی به شکل حلقویمیباشد.کپسید
پوشش غیر پروتئینی
وکربوهیدراتها است پوشیده شده است. ویروسهای ناقص Defctive Virus
تشکیل شدهاند، ولی بدون ویروس کمکی توان تکثیر ندارند. که به این ویروس کمکی Helper ویروس گفته میشود. ویروسهای ناقص در ساختمان ژنتیکی خود نقصی دارند و در خلال تکثیر در داخل سلول بوجود میآیند و چون این ویروسها میتوانند تکثیر ویروسهای معمولی را مختل کنند تصور میشود که این ویروسها با تکثیر زیاد خود از تکثیر ویروسهای معمولی جلوگیری میکنند پس در بهبود بیماری نقش دارند.ویریون
سودو ویریون
ویروتید
ویروسوئید

ویروسهای گیاهی
،قارچها ، گلسنگها ،خزهها ، سرخسها و گیاهان عالی دیده شدهاند. ولی در گیاهان عالی بیش از گیاهان پست مورد مطالعه قرار گرفتهاند. ویروسها به گیاهان زراعی خسارت عمدهای وارد میسازند. چون پارهای از ویروسهای گیاهی چندان شباهتی با ویروسهای دیگر ندارند بنابراین گروه مستقلی را تشکیل میدهند. ولی بعضی از آنها دارای خصوصیات مشترک بوده و میتوان آنها را در یک گروه قرار داد. این گروهها به شرح زیر هستند. - ویروسهای میلهای یا رشتهای
- ویروسهای ایزو دیامتریک
- ویروسهای باسیلی شکل
- ویروئیدها: بیماریزاهایی شبیه ویروسها هستند که در میزبان خود نوکلئو پروتئین تولید نمیکنند.

ویروسهای جانوری
ویروس از انواع مختلف جانوران از تک یاختگان تا انسان جدا شده است. میزبان مهم ویروسها در بیمهرهگان ، بندپایان هستند خصوصا کنهها و حشرات. پارهای از ویروسها در عین حال که در حشرات تکثیر مییابند میتوانند در گیاه یا در جانور مولد بیماری باشند، ولی برای خود حشرات بیماریزا محسوب نمیشوند. ویروسها در اکثر مهرهدارانفعالیت دارند و در ماهیها ، دوزیستان ، پرندگان و پستانداران بیماریهایی تولید میکنند که گاهی علایم آنها به صورت تومور نمایان میشود. ویروسها در انسان نیز بیماریهای گوناگونی مانند اوریون ، سرخک ، تب زرد ، آبله ، آنفلوانزا و ... ایجاد میکنند.
تکثیر ویروسها
اسید نوکلئیک هر ویریون فقط تعداد معدودی از ژنهای لازم برای سنتز ویروسهای جدید را دارا میباشد. اکثر آنزیمهای ویروسها توسط سلول میزبان ساخته میشوند. نقش آنزیمهای ویروس تقریبا بطور کامل با همانند سازی و آماده کردن اسید نوکلئیک ویروسی ارتباط دارد و هرگز با دستگاه سنتز پروتئینی را تولید انرژی رابطهای ندارد. مراحل 5 گانه تکثیر ویروس در سلول میزبان به صورت زیر است.
- مرحله رونشینی ویروسها بر روی سلول
- مرحله ورود و نفوذ در سلول
- مرحله بیوسنتز اجزای ویروسی
- مرحله رسیدن و کامل شدن ویروس
- مرحله آزاد شدن ویروس از سلول میزبان و نفوذ آن در سلولهای سالم
شیمی درمانی علیه ویروسها
داروهایی که در مراحل مختلف تکثیر ویروسها در بدن میزبان اثر میکنند در تجربیات آزمایشگاهی موثر شناخته شدهاند. ولی از نظر بالینی آمانتادین ، آسیکلوویر ، ویدارابین وتیو سمی کاربازون مفید شناخته شدهاند. در اغلب بیماریهای ویروسی تکثیر ویروس تقریبا قبل از ظاهر شدن علایم بیماری پایان پذیرفته است. مساله دیگر پیدایش ویروسهای جهش یافته مقاوم نسبت به این داروها میباشد و کثرت وقوع آنها به اندازه باکتریها میباشد.شیمی درمانی علیه ویروسها در مراحل اولیه است و میتوان در آینده داروهایی علیه ویروسها کشف کرد.
سلولهای بنیادی
سلولهای بنیادی
 تلاش برای استفاده از سلولهای بنیادی جنینی از حدود 20 سال پیش با کار بر روی حیوانات به ویژه موشهای آزمایشگاهی شروع شد. در طی این سالها، آزمایشات زیادی در جهت تبدیل سلولهای بنیادی جنینی موش به انواع سلولها و پیوند زدن آنها صورت گرفت که به موفقیتهای قابلتوجهی انجامید. در جوار این موضوع، سلولهای بنیادی انسان نیز مورد توجه قرار گرفت تا اینکه بالاخره در سال 1998 اولین گزارش موفقیتآمیز از تکثیر و تمایز سلولهای بنیادی جنینی انسان در آمریکا منتشر شد. اما با توجه به بروز برخی محدودیتها در تولید و استفاده از سلولهای بنیادی جنینی (که تلاش برای رفع آنها ادامه دارد) در چند سال اخیر، موج جدیدی از تحقیقات بر روی سلولهای بنیادی بالغ شروع شد که کماکان ادامه دارد.
تلاش برای استفاده از سلولهای بنیادی جنینی از حدود 20 سال پیش با کار بر روی حیوانات به ویژه موشهای آزمایشگاهی شروع شد. در طی این سالها، آزمایشات زیادی در جهت تبدیل سلولهای بنیادی جنینی موش به انواع سلولها و پیوند زدن آنها صورت گرفت که به موفقیتهای قابلتوجهی انجامید. در جوار این موضوع، سلولهای بنیادی انسان نیز مورد توجه قرار گرفت تا اینکه بالاخره در سال 1998 اولین گزارش موفقیتآمیز از تکثیر و تمایز سلولهای بنیادی جنینی انسان در آمریکا منتشر شد. اما با توجه به بروز برخی محدودیتها در تولید و استفاده از سلولهای بنیادی جنینی (که تلاش برای رفع آنها ادامه دارد) در چند سال اخیر، موج جدیدی از تحقیقات بر روی سلولهای بنیادی بالغ شروع شد که کماکان ادامه دارد.ایران بهعنوان یکی از معدود کشورهای تولیدکنندة سلولهای بنیادی جنینی
فناوری تولید و پرورش سلولهای بنیادی جنینی در دنیا کار جدیدی است؛ بهطوریکه پس از کشف سلولهای بنیادی جنینی موش در سال 1981، اولین سلولهای بنیادی جنینی انسان در سال 1998 تکثیر شد. در این میان، پس از چند کشور پیشرفته نظیر آمریکا، استرالیا، اسرائیل، سنگاپور، انگلستان، ژاپن، سوئد، هند و کرهجنوبی که به فناوری تکثیر و پرورش این سلولها دست پیدا کردهاند، ایران از جمله معدود کشورهایی است که به این مهم دست یافته است و لذا فاصله کشورمان در این مورد از دیگر کشورهای پیشرو چندان زیاد نیست.
منشأ سلولهای بنیادی بالغ
سلولهای بنیادی بالغ همانطور که از نامشان مشخص است، پس از تولد از فرد گرفته میشوند. برای مثال این سلولها را میتوان از بافت مغز استخوان یک فرد سالم تهیه کرد. البته بر اساس یافتههای اخیر، برخی معتقدند که هر بافتی دارای سلولهای بنیادی خاص خود است. بهطور مثال، مشخص شده که قلب، مغز و ماهیچههای اسکلتی هر کدام دارای سلولهای بنیادی خاص خود هستند و همة این سلولها در بدن یک فرد بالغ وجود دارند. بهعنوان مثال، سلولهای بنیادی قلبی بیشتر در ناحیه اپیکس (Apex) قلب و سلولهای بنیادی مغزی عمدتاً در دیوارة بطن مغز متمرکز هستند. با این حال دقیقاً مشخص نیست که منشأ این سلولهای بنیادی گوناگون، چه سلولی است و آیا منشأ همه اینها همان سلولهای مغز استخوان هستند که هر یک به سمت اندام خاصی مهاجرت کرده و به سلولهای بنیادی خاص آن تبدیل میشوند، یا منشأ دیگری برای آنها وجود دارد.
منشأ سلولهای بنیادی جنینی
 بنیاختههای جنینی در مرحله بلاستوسیست از تودة سلولی داخلی یا Inner Cell Mass گرفته میشوند. بلاستوسیست یکی از مراحل دوران جنینی است که به لحاظ مرفولوژی، شبیه یک توپ توخالی است.
بنیاختههای جنینی در مرحله بلاستوسیست از تودة سلولی داخلی یا Inner Cell Mass گرفته میشوند. بلاستوسیست یکی از مراحل دوران جنینی است که به لحاظ مرفولوژی، شبیه یک توپ توخالی است.
سلولهای محیط این توپ تروفوبلاست (Trophoblast) هستند که جفت را میسازند. در داخل این توپ هم تعدادی سلول جمع شدهاند که در مراحل بعدی، به جنین تبدیل میشوند. اگر این تودة سلولهای داخلی را برداشته و در محیط آزمایشگاهی کشت بدهند، بنیاختههای جنینی ایجاد میشوند. اما هنوز دقیقاً مشخص نیست که آیا این توده سلولهای داخلی منشأ بنیاختههای جنینی هستند، یا اینکه فرآیند مذکور، حاصل شرایط محیطی بوده و تودة سلولهای داخلی در محیط آزمایشگاهی سلولهای دیگری را میسازند که آنها به بنیاخته جنینی بدل میشوند.
تلومر
کارکرد و ویژگی های تلومر
 انتهای مولکول خطی که بطور معمول چسبنده میباشد با حضور تلومر و ساختمان آن این چسبندگی را از دست میدهد. از این رو تلومر مانند سپری کروموزم را از ایجاد پیوستگیهای نابجا و غیر صحیح و تخریب بهوسیله آنزیمهای اگزونوکلئاز سلول محافظت مینماید. همچنین، مشخص شده است که تلومر درمکان یابی و جایگیری کروموزم در هسته و خاموشی انتخابی ژنهای مجاور خود، ایفای نقش میکند علاوه برا ین، تلومر نقش اساسی دیگری در ابتدای سنتز خود ایفا میکند که در ارتباط با رونویسی میباشد. در واقع در انتهای یک DNA خطی کار آنزیم های همانند ساز در رشته پیرو به مشکل برخورد میکند چرا که این آنزیمها برای برداشتن آخرین پرایمر و قراردادن آخرین بازها، پایانه 'OH۳- را در اختیار ندارند. آنزیم تلومراز (Telomerase) این مشکل را با ستنز تلومر حل میکند.
انتهای مولکول خطی که بطور معمول چسبنده میباشد با حضور تلومر و ساختمان آن این چسبندگی را از دست میدهد. از این رو تلومر مانند سپری کروموزم را از ایجاد پیوستگیهای نابجا و غیر صحیح و تخریب بهوسیله آنزیمهای اگزونوکلئاز سلول محافظت مینماید. همچنین، مشخص شده است که تلومر درمکان یابی و جایگیری کروموزم در هسته و خاموشی انتخابی ژنهای مجاور خود، ایفای نقش میکند علاوه برا ین، تلومر نقش اساسی دیگری در ابتدای سنتز خود ایفا میکند که در ارتباط با رونویسی میباشد. در واقع در انتهای یک DNA خطی کار آنزیم های همانند ساز در رشته پیرو به مشکل برخورد میکند چرا که این آنزیمها برای برداشتن آخرین پرایمر و قراردادن آخرین بازها، پایانه 'OH۳- را در اختیار ندارند. آنزیم تلومراز (Telomerase) این مشکل را با ستنز تلومر حل میکند.
|
با توجه به اینکه در هر دور همانند سازی با فعالیت تلومر به طول تلومر افزوده میشود به نظر میرسد که اندازه تلومر پیوسته افزایش مییابد اما درواقع چنین مسئلهای رخ نمیدهد بلکه حتی در سلولهای سوماتیک جانداران پر سلولی مثل انسان طول تلومر پیوسته کاهش مییابد. این سلولها طی تمایز، توانائی تولید آنزیم تلومراز را از دست دادهاند بنابراین در هر دور همانند سازی در این سلولها آنزیمهای درگیر درانتهای تلومر با مشکل عدم وجود پایانه روبرو شده وقسمتی از انتهای تلومر همانند سازی نمیگردد. این مسئله طی تقسیمات متوالی باعث کاهش تدریجی طول تلومر میگردد.
 البته سلولهای سوماتیکی که به سرعت تقسیم میشوند و سلولهای تولید مثلیٍ، توانائی تولیدمثل تلومراز را دارند وبنابراین دچار این کاهش در طول تلومر نمیگردد.
البته سلولهای سوماتیکی که به سرعت تقسیم میشوند و سلولهای تولید مثلیٍ، توانائی تولیدمثل تلومراز را دارند وبنابراین دچار این کاهش در طول تلومر نمیگردد.علاوه بر آنچه بالا گفته شد احتمال وجود سیستم کاهش طول وابسته به پروتئین که به اصطلاح باعث فرسایش یا خوردگی تلومر میشود نیز مطرح شده است.
مشاهدات نشان داده است که در بین سلولهای سوماتیک سلولهای عصبی در رابطه با طول تلومر استثنا میباشند. چراکه در این سلولها طول تلومر همواره تقریباٌ ثابت باقی میماند. باتوجه به اینکه این سلولها پس ازدوران جنینی بطور معمول تقسیم نمیشوند ثبات طول تلومر در آنها مکانیزم کاهش طول تلومر براساس همانند سازی ناقص و یا هر مکانیزم دیگر را که وابسته به تقسیم سلولی باشد تائید میکند.
سوالی که در اینجا مطرح میگردد این است که آیا در سلولهایی که بطور مداوم تقسیم میشوند مثل سلولهای تولید مثلی و یا تک سلولیهای یوکاریوتیک که در آنها تقسیم سلولی به معنای تولید نسل است و بدون محدودیت دنبال میشود، طول تلومر به علت وجود آنزیم تلومراز پیوسته افزایش مییابد؟
مشاهدات و آزمایشات انجام شده بر روی سلولهای مخمر نشان دادهاند که نوعی تعادل بین کاهش وافزایش طول تلومر در این سلولها وجود دارد. بدین صورت که سیستمهای مولکولی خاصی با کاهش تدریجی طول تلومر و رسیدن آن به یک آستانه معین امکان افزایش طول آنرا فراهم میآورند.
به عنوان مثال پیشنهاد شده است که پروتئین متصل شونده به تلومر به نامTelomere Binding Protein دارای تعدادی جایگاه اتصال روی تلومر است ،هرگاه که این پروتئین به تعداد معین ( و یا بیشتر از آن ) در اتصال با تلومر وجود داشته باشد تلومراز امکان اتصال و سنتز دنباله تلومر را نمییابد. اما وقتی به علت کاهش طول تلومر، چه با فرسایش و چه با همانند سازی ناقص، تعداد جایگاههای TBPو در نتیجه تعداد مولکولهای این پروتئین بر روی تلومر کاهش پیدا کند، تلومراز اجازه مییابد به تلومر متصل شده و آنرا طویل کند این طویل شدن باعث ایجاد جایگاههای جدید برای اتصال تعدادی از مولکولهای این پروتئین به تلومر میشود که این امر دوباره باعث جلوگیری از افزایش طول مجدد
تلومر توسط تلومراز شده و تعادل بین کاهش وافزایش طول حفظ میشود.
همانطورتلومر و طول عمر که گفته شد وجود تلومر به عنوان سپر حفاظتی برای محافظت از ژنوم سلول یوکاریوتی اهمیت حیاتی دارد وکاهش زیاد طول تلومر منجر به از بین رفتن توانائی عملکرد این ساختار در انجام وظایف خود شده ودرنهایت سلول را به سوی نابودی میبرد. مشاهدات متعدد نشان دادهاند که سلولهای سوماتیک انسانی طبیعی، که در سیستم در شیشه(in vitro)کشت داده شدهاند ،تنها میتوانند تعداد محدودی تقسیم را انجام دهد و پس از آن رشد آنها متوقف شده و سلولها دچار سالخوردگی میشوند پس از اینکه کاهش طول تلومر به حد بحرانی برسد فرکانس بالائی از نوترکیبیهای کروموزمی مشاهده میشود همین امر میتواند عامل سالخوردگی ونهایتاٌ نابودی سلول گردد. این اتفاق دربدن موجودات زنده (in vivo) نیز رخ میدهد و تحقیقات ارتباط طول عمر موجودات زنده پرسلولی وکاهش طول تلومر را نشان میدهند. به عنوان مثال در یک بررسی بر روی Rat مشاهده شد که کاهش طول عمر تلومر در بافتهای سوماتیک این جانور در جنس نر بیشتر ( سریعتر ) از جنس ماده است.و این مطلب با طول عمر آنها که در مادهها بیش از نرهاست مطابقت دارد.
همین مسئله باعث شد که بحثهائی پیرامون افزایش مدت عمر بشر وحتی جاودانگی بشر مطرح گردد و دانشمندان در تلاش هستند که ابتدا اینکار را با ساختن حیوانات آزمایشگاهی مثلاٌ موشهائی با عمرهای طولانی تر ازحد معمول به مرحله عمل برسانند.

تلومر و سرطان
برخلاف سلولهای سوماتیک طبیعی، سلولهای سرطانی میتوانند بطورمتوالی تقسیم شده و خطوط سلولی نامحدود تولید نمایند. ( مثل سلولهای هلا) برای داشتن چنین خصوصیتی، این سلولها باید توانائی حفظ طول تلومر خود را داشته باشند. این سلولها میتوانند این توانائی را با تولید آنزیم تلومراز بدست آوردند. در واقع ایجاد توانائی تولید این آنزیم که میتواند توسط ویروسها و یا سایر عوامل جهش زا، در سلولهای سوماتیک بدن ایجاد گردد یکی از عوامل سرطانی شدن این سلولها بشمار میرود.
از سوی دیگر همین مسئله از سوی پژوهشگران به عنوان پاشنه آشیلی برای سلولهای سرطانی تلقی میشود. چراکه با طراحی درمانهائی که ساز و کار حفظ تلومر را در این سلولها هدف قرار میدهد میتوان این سلولهای نامیرا را به سلولهایی با تقسیمات محدود و درواقع میرا تبدیل کرده و نابود نمود.
یکی از راحترین راهها برای انجام اینکار هدف قراردادن تلومراز است. چراکه این آنزیم مسئول نامیرائی در سلولهای سرطانی است. اما از سوی دیگر مشخص شده است که گاهی چنین مباررزهای تأثیر معکوس میدهد به عنوان مثال در یک مطالعه درموشهای آزمایشگاهی مشاهده شده است که این نوع درمان اگر چه باعث کاهش توان حیاتی سلولهای سرطانی میشود اما فراوانی لنفوما را در این موشها افزایش میدهد. این امر ممکن است به علت افزایش امکان ایجاد نابسامانیهای کروموزمی به علت کاهش طول تلومرها باشد به بیان دیگر در این روش امکان بوجود آمدن خطوط سلولی جدید که نسبت به درمان مقاومت نشان میدهند وجود دارد.
ایراد دیگر این روش این است که پس از تحت تأثیر قرار دادن آنزیم تلومراز ( که به طرق مختلف مثلاٌ طراحی شناساگرهای مکمل برای بخش RNA این ریبونوکلئوپروتئین که کار آن را مختل میکند،امکانپذیر است) بایدمنتظر ماند تا عوامل کاهش طول تلومر به تدریج اندازه تلومرها را کاهش دهند و این امر مستلزم سپری شدن چندین مرحله تقسیم سلولی است. بنابراین، درمانهای مبتنی بر توقف فعالیت تلومراز نمیتوانند بطور مستقیم وبی واسطه بر سلولهای سرطانی تأثیر گذار باشند وراه بهتر برای انجام درمان براساس تلومر هدف قراردادن پروتئینهای شرکت کننده در ساختمان آن برای از هم پاشاندن این ساختار و یا هدف قراردادن پروتئینهای شرکت کننده در مسیرها و واکنشهای منتهی به ایجاد ساختار و یا هدف قراردادن پروتئینهای آن میباشد.که این امر خود مستلزم شناخت دقیقتر از ساختمان تلومر و چگونگی تشکیل آن با انجام پژوهشهای بیشتر در این زمینه است.
میتوکندری

نام میتوکُندری ترکیبی است از دو واژه یونانی Mito به معنای رشته وchondrion به معنی دانه. چون ایناندامک اغلب رشتهای یا به صورت دانههای کوچک در سیتوپلاسم همه سلولهای یوکاریوتی وجود دارد.میتوکندریها در تمام یاختههای دارای تنفس هوازی به جز در باکتریها که آنزیمهای تنفسی آنها در غشای سیتوپلاسمی جایگزین شدهاند وجود دارند. این اندامکها، نوعی دستگاه انتقال انرژی هستند که موجب میشوند انرژی شیمیایی موجود در مواد غذایی با عمل فسفوریلاسیون اکسیداتیو، به صورت پیوندهای پرانرژی فسفاتATP) ذخیره شود. 
تاریخچه
اولین بررسیهای انجام شده بر روی میتوکندریها، در سال 1894 بهوسیله آلتمن صورت گرفت که آنها را بیوپلاست یا جایگاههای زنده نامید. و نظر داد که بین واکنشهای اکسایش و کاهش سلول و میتوکندری وابستگی وجود دارد. در سال (1897) بتدا با بررسیهای بیشتر آنها را میتوکندری نامید و در 1900، میکائیلیس به کمک معرف رنگی سبز ژانوس میتوکندری را در سلولهای زنده مشاهده کرد. واربورگ در سال 1913 آنزیمهای تنفسی را در این اندامک نشان داد. سرانجام برای اولین بار، در سال 1934، بنسلی و هر، توانستند آنها را از سلولهای کبدی جدا کرده و بعد آن بررسیهای بیشتر و عملیتر روی آن صورت گرفت.
تالاسمی
تالاسمی
هموگلوبین
هموگلوبین مولکول اصلی داخل گویچههای قرمز است که از هم و زنجیرههای پروتئنی یا گلوبین تشکیل شده است. در هر زنجیره گلوبین یک مولکول هم وجود دارد که اکسیژن را توسط آهن خود حمل میکند. پس تولید هموگلوبین نیاز به به تامین آهن و ساخت هموگلوبین دارد. بر اساس نوع زنجیره پروتئینی چند نوع هموگلوبین وجود دارد: هموگلوبین A: هموگلوبین طبیعی در بالغین عمدتا همولگوبین A میباشد که تقریبا حدود 98% از هموگلوبین جریان خون را تشکیل میدهد و از زنجیره 4 تایی حاوی دو زنجیره آلفا و دو زنجیره بتا ساخته میشود. (α2β2)
انگشت نگاری ژنتیکی
انگشت نگاری ژنتیکی
 فرض کنید که در یک زندان به علت مظنون بودنتان به یک جرم یا تبهکاری محبوس شده اید و هیچگونه مدرکی دال بر بی گناهی شما وجود ندارد، روزهای سرد وسیاه حبس تا آخر عمر انتظارتات را می کشد تا اینکه با کمک یک آزمون علمی شگرفت انگیز بی گناهی شما ثابت می گردد و ناباورانه آزاد می شوید. این تصور خیالی دقیقا با بخشی از واقعیت زندگی ویلیام توماس از ایالت کنتاکی عجین شده است فردی که در سال 1993 در جریان یک بزهکاری مظنون شناخته شد اما همواره در دادگاه بر بی گناهی خود پافشاری نمود تا اینکه 6 سال بعد بی گناهی وی با آزمون DNA به اثبات رسید. تکنیک انگشت نگاری ژنتیکی، در سال 1985 توسط الک جفری در دانشگاه لاسیستر ابداع شد. او سرگرم مطالعه برروی ژن میوگلوبین بود. پروتئین حاصل از این ژن موجب ذخیره اکسیژن در ماهیچه می شود.جفری در ضمن تحقیق خود در یافت که نواحی از ژن میوگلوبین فاقد اطلاعات لازم برای ساخت میوگلوبین می باشد و نقشی در سنتز پروتئین میوگلوبین ندارد. این نواحی از ژن شامل نواحی غیر عادی از لحاظ بازهای نوکلئوتیدی بود که در آن این بازها، چندین بارپشت سر هم تکرارمی شدند. جفریس این توالیها را ماهوارک نامید او اندیشید که ممکن است این توالیها سر نخی برای دانشمندان ژنتیک باشد تا گره از بعضی معماهای ژنوم بگشایند. او این قطعات حاوی بازهای نوکلئوتیدی تکراری را پس ازجداسازی ازکل ژنوم وارد باکتری نمود و بااستفاده ازتکنیک نشاندار کردن قطعات حاصل از تکثیرو الکتروفورز، الگوهای مختلف باندی را در روی ژل مشاهده کرد. این دانشمند برجسته مشاهده کرد که وجود توالیهای بازی تکراری تنها مختص ژن میوگلوبین نیست بلکه در سایر نقاط DNA ژنومی نیز این توالیها را می توان یافت. عجیبتر اینکه در هر فرد این توالیها منحصر به آن فرد است از والدین به نسبت مساوی به فرزندان منتقل می شود.
فرض کنید که در یک زندان به علت مظنون بودنتان به یک جرم یا تبهکاری محبوس شده اید و هیچگونه مدرکی دال بر بی گناهی شما وجود ندارد، روزهای سرد وسیاه حبس تا آخر عمر انتظارتات را می کشد تا اینکه با کمک یک آزمون علمی شگرفت انگیز بی گناهی شما ثابت می گردد و ناباورانه آزاد می شوید. این تصور خیالی دقیقا با بخشی از واقعیت زندگی ویلیام توماس از ایالت کنتاکی عجین شده است فردی که در سال 1993 در جریان یک بزهکاری مظنون شناخته شد اما همواره در دادگاه بر بی گناهی خود پافشاری نمود تا اینکه 6 سال بعد بی گناهی وی با آزمون DNA به اثبات رسید. تکنیک انگشت نگاری ژنتیکی، در سال 1985 توسط الک جفری در دانشگاه لاسیستر ابداع شد. او سرگرم مطالعه برروی ژن میوگلوبین بود. پروتئین حاصل از این ژن موجب ذخیره اکسیژن در ماهیچه می شود.جفری در ضمن تحقیق خود در یافت که نواحی از ژن میوگلوبین فاقد اطلاعات لازم برای ساخت میوگلوبین می باشد و نقشی در سنتز پروتئین میوگلوبین ندارد. این نواحی از ژن شامل نواحی غیر عادی از لحاظ بازهای نوکلئوتیدی بود که در آن این بازها، چندین بارپشت سر هم تکرارمی شدند. جفریس این توالیها را ماهوارک نامید او اندیشید که ممکن است این توالیها سر نخی برای دانشمندان ژنتیک باشد تا گره از بعضی معماهای ژنوم بگشایند. او این قطعات حاوی بازهای نوکلئوتیدی تکراری را پس ازجداسازی ازکل ژنوم وارد باکتری نمود و بااستفاده ازتکنیک نشاندار کردن قطعات حاصل از تکثیرو الکتروفورز، الگوهای مختلف باندی را در روی ژل مشاهده کرد. این دانشمند برجسته مشاهده کرد که وجود توالیهای بازی تکراری تنها مختص ژن میوگلوبین نیست بلکه در سایر نقاط DNA ژنومی نیز این توالیها را می توان یافت. عجیبتر اینکه در هر فرد این توالیها منحصر به آن فرد است از والدین به نسبت مساوی به فرزندان منتقل می شود.
امروزه واژه میکروساتلایت و مینی ساتلایت واژه بسیار آشنا برای متخصصان علم ژنتیک می باشند .معمولترین تکرارهای موجود در ژنوم پستانداران،تکرارهای(CA)n و(CT)n و (dG-dT) می باشند. تعداد بازهای موجود در هر ماهوارک ممکن است به 2،3، 4 و حتی بیشتر برسد که طبق قاعده خاصی در ژنوم تکرار می شود. برای چنین آزمونهای نیاز به بررسی تمام ژنوم نمی باشد و همانطور که وجود یک خال در بدن می تواند دلیلی محکم بر شناسائی یک بی گناه یا قاتل باشد بررسی یک ناحیه کوچک از ژنوم فرد مظنون نیز چنین مدارکی ارزشمندی را مهیا خواهد نمود. این مقاله سعی دارد که در یک نگرش اجمالی، بعضی از کاربردهای انگشت نگاری ژنتیکی را همراه با وقایع شگفت انگیزی که این تکنیک در آنها مورد استفاده قرار گرفته است ، را بیان نمایید.
آزمون انساب (تعیین هویت): در ژوئیه 1997، مونیکالمود، زنی که 8 ماهه آبستن بود به همراه همسر خود توسط نیروهای امنیتی ایالت بوئنوس آیرس در کشور آژانتین ربوده شد. سرنوشت این زوج و کودکی که مونیکا در رحم داشت ناشناخته باقی ماند. بعدها مردی که از یک اردوگاه سری در آژانتین آزاد شده بود به مادر بزرگ مونتیکا گفت که دختر او در اردوگاه ویژه امنیتی زندانی شده بود. در سال 1985 در همان اردگاه زنی که سابقا پلیس بود، ادعا کرد که والد قانونی دختری است که در زندان متولد شده است . دو سال بعد آزمایش انگشت نگاری ژنتیکی با قطعیت 98/99 درصد دختر را متعلق به مونیکا دانست و این کودک به مادر بزرگ خانواده بازگردانده شد. از آن پس تکنیک انگشت نگاری ژنتیکی در حل مشاجرات مربوط به تعیین والدین حقیقی کودکان استفاده شد.
شناسائی مجرمان : در مواردی که قربانی در اثر تجاوز جنسی کشته شده باشد، می توان با تهیه نمونه سلولهای جنسی موجود در مهبل قربانی و همچنین تهیه نمونه خون یا بافت از مقتول، سرنخی برای تشخیص قاتل به دست آورد. در این روش پس از استخراج DNA از نمونه ها و همچنین مقایسه الگوهای باندی حاصل از نمونه خون مقتول و با الگوهای باندی حاصل از نمونه حاوی مخلوطی از سلولهای جنسی مقتول و قاتل ، الگوهای باندی متعلق به قاتل را تشخیص داد. انگشت نگاری ژنتیکی همچنین می تواند برای شناسائی اجسادی که تخریب و متلاشی شده اند و یا به هر علتی قابل تشخیص نیستند ،مورد استفاده قرار گیرند. در سال 1989 پلیس بقایای جسدی را که در یک فرش پیچیده شده بود را پیدا کرد، باز سازی چهره جسد، پلیس را مشکوک نمود که جسد متعلق به کارنی پرایس نوجوانی است که در سال 1981 ناپدید شده است . اما هیچ کس نمی توانست مطمئنن باشد. اریکاهاگلبرگ مقداری اندکی DNA از یکی از استخوانهای جسد، استخراج کرد و آن را برای تجزیه و تحلیل به آزمایشگاه چفریس فرستاد.مدارک مستدل بر پایه انگشت نگاری ژنتیک نشان داد که جسد واقعا متعلق به کارنی پرایس است و این مدرک توسط دادگاه بریتانیایی وقت، پذیرفته شد. این اولین واقعه بود که در آن انگشت نگاری DNA به عنوان مدرکی در دادگاه پذیرفته شد .
تشخیص سرطانها : در برخی از انواع سرطانها ، به طور قابل توجهی افزایش یا کاهش تعداد توالیهای میکروساتلایت دیده می شود.چنین تغییراتی باعث ایجاد ریز ماهوارکهای متفاوت می شود که براحتی قابل تشخیص هستند.
برآورد پارامترهای جمعیتی : با استفاده از انگشت نگاری DNA ، تمایز ژنتیکی ، تعداد مهاجران در هر نسل ، میزان همخونی را برآورد نمود به خصوص با استفاده از این ابزار قدرتمند، زمان اشتقاق گونه ها را نیز می توان محاسبه نمود.
تایید صحت ادعاهای شبیه سازی: یکی از موارد جالب کاربرد انگشت نگاری ژنتیکی، آزمون صخت ادعاهای موجود در رابط با شبیه سازی می باشد. به عنوان مثال در گوسفند دالی که اولین پستانداری بود که به روش انتقال هسته سوماتیک بوجود آمد این آزمون توسط Ashworth و همکاران در سال 1998 صورت گرفت و با انگشت نگاری ژنتیکی مشخص شد که او حامل 7 اللی ریزه ماهوارهای است که در جمعیت سلولی سازنده اش وجود داشته است و تردیدهای موجود بی مورد است.
مطالعات مربوط به حفاظت گونه ها و رفتار شناسی گونه های در حال انقراض: تکنیک انگشت نگاری ژنتیکی همچنین توسط محققان دانشگاه کالیفرنیا بعنوان ابزاری برای ردیابی رفتار آمیزشی گروهی از شمپانزه های وحشی در جنگلی انبوه استفاده شد. این محققان موهای باقیمانده موجود در آشیانه های موقت موجود در نوک درختانی که شمپانزه های در آن می خوابیدند را جمع آوری نمودند و DNA مورد نیاز را از ریشه موها استخراج کردند، با مقایسه الگوهای باندی ماده ها و نرهای بالغ و 13 بچه شمپانزه در یافتند که 7 بچه متعلق به نرهای این گروه کوچک نیست . بنابراین این نظریه قوت یافت که در طی شب حداقل بعضی از شمپانزه های ماده می بایستی به جنگلهای مجاور رفته و با نرهایی از گروهای دیگر برخورد نموده اند. کشف چنین روابطی با انگشت نگاری ژنتیکی ، می تواند توضیح دهد که چگونه در گروهای کوچک شمپانزه ها نیز حفظ تنوع ژنتیکی و اجتناب از همخونی امکان پذیر می باشد.
آپوپتوسیس
آپوپتوسیس
در مراحل مشخصی از تکامل یک ارگانیسم چند سلولی ، بعضی از سلولها باید بمیرند. این فرآیند که به خوبی برنامه ریزی شده است آپوپتوسیس (مرگ برنامه ریزس شده سلولی) نام دارد. اهمیت بیولوژیکی این پدیده اولین بار در مطالعاتی که بر روی یک کرم کوچک ، C.elegans که یک نماتد است انجام شد.اگر آپوپتوسیس اتفاق نیفتد ممکن است نقصهای تکاملی یا سرطان اتفاق بیفتند. آپوپتوسیس در مسیر خود بوسیله بسیاری از پروتئینها کنترل می شود ، که می توانند منجر به شروع آپوپتوسیس یا جلوگیری از آن شوند. آپوپتوسیس می تواند بوسیله محرکهای گوناگون خارج سلولی (مسیر خارجی) یا از داخل سلول (مسیر داخلی) تحریک شود. محرکهای خارجی ممکن است اشعه ، باز گرفتن یک فاکتور رشد و یا گلوکوکرتیکوئید باشد. یک محرک داخلی ممکن است آسیب خودبخودیی در DNAسلول باشد.اولین علامت قابل مشاهده آپپتوسیس متراکم شدن کروماتین و چروک شدن سلول می باشد.غشای سلول چروک می شود (غشا برامده می شود) ، و سلول شروع به جدا شدن می کند( هسته قطعه قطعه می شود ، DNA تکه تکه می شود).و بقایای بدنه آپوپتوز شده سلول ، در فرایندی که لیز نام دارد حل می شوند.
تنظیم آپپتوسیس
آنزیمهای کسپاز caspase (که آبشار یا کاسپاز نیز ترجمه شده) خانواده ای از پروتئازهای آسپارتات می باشند که دارای سیستئین هستند و در آپپتوسیس (مرگ برنامه ریزی شده سلولی ) نقش اصلی را ایفا می کنند.آنها همدیگر رادر یک توالی تعریف شده فعال و غیر فعال می کنند . اتصال یک لیگاند ، Fas ، مربوط به سلولهای T سایتوتوکسیک) cytotoxic (Tcellبه رسپتورFas (که CD95 نیز نامیده می شود) باعث فعال شدن داخل سلولی پروتئین آداپتور یا FADD (دومین مرگ وابسته به Fas) می شود. بدنبال این اتصالات ، فعال شدن پیش کسپاز 8 (procaspase 8 ) منجر به فعال شدن کسپاز 8 (caspase 8 ) می شود. کسپاز 8 باعث آزاد شدن سیتوکروم C از میتوکندری و فعال شدن چندین کسپاز سلولی متفاوت دیگر می شود. در مورد کسپازها دو مسیر وجود دارد در مورد سلولهای نوع I (در تیموسیتها و فیبروبلاستها) ، کسپاز 8 مستقیما باعث فعال شدن کسپاز 3 می شود. در سلولهای نوع II ، از قبیل هپاتوسیتها ، کسپاز 8 باعث جدا شدن Bid که عضوی از خانواده Bcl-2 است می شود.
ژنوم انسان و موش دارای 13 ژن برای کسپاز (caspase) است (ژنهای 1-12 و 14). کسپازهای 3 و 6-10 انسان در آپپتوسیس نقش دارند ، و بقیه در التهاب مؤثرند.مسیر متداول مرگ سلولی از طریق مرگ سلولی برنامه ریزی شده بوسیله p53 هدایت می شود . پروتئین p53 در چرخه سلولی در مناطق کنترل وضعیت (check point) در صورت وجود جهش یا آسیب در DNA باعث ممانعت از تکثیر سلول یا مرگ سلول می شوند. در بیش از 50 درصد سرطانهای دنیا جهش در پروتئین p53 یافت شده است.از این رو p53 از جمله مهمترین اهداف ژن درمانی تومور به حساب می آید. از دیگر تنظیم کننده های آپوپتوسیس اعضاء خانواده bcl-2 می باشند.(اسم bcl از سلولهای B لنفوما آمده که در آن یک تومور بدخیم از لنفوسیتهای B منشا می گیرد که دلیل آن وجود جهش در ژن آن می باشد.
ژن bcl-2 یک پروتوانکوژن است و سلولها را از مرگ سلولی محافظت می کند.(پروتوانکوژن ها : ژنهای معمولی سلولی مرتبط با رشد , تکثیر , تمایز و فعالسازی عمل نسخه برداری می باشند.) در تعدادی از ویروسها , خانواده ای از پروتئینها به نام "مشابه bcl-2" وجود دارند که به ویروس توانایی جلوگیری از مرگ سلولی اعطا می کند این امر به زنده ماندن سلول و استفاده بیشتر ویروس از آن منجر می شود. پروتئینهای دیگری همچون Bax در بقای سلول نقش دارند که در موارد تکثیر بیش از حد سلول می توان از تغییر وضعیت ژن آنها برای درمان تومور استفاده نمود.از سوی دیگر آنزیم ICE (آنزیم تغییر دهنده انترلوکین یک) باعث پیشرفت مرگ سلولی می شود. می بینیم که عوامل متعددی در آپپتوسیس نقش دارند که چرخه پیچیده ای از ارتباط بین مولکولها را ایجاد کرده اند.



